Som art har vi mennesker udviklet os side om side med billioner af bakterier, der har dannet et helt økosystem i mundhulen, i mave-tarm-kanalen og på huden (1). Af alle de celler, der udgør dig — ét enkelt Homo sapiens — anslås omkring 76 % at være menneskeceller; resten er bakterieceller, og sammen udgør de dit "udvidede genom" (2). De billioner af mikrober i og på din krop udtrykker 100 gange så mange gener som det menneskelige genom alene (2). Funktionen af dette udvidede genom anerkendes i stigende grad som en vigtig medspiller for helbredet — og den funktion er tydeligt påvirket af kosten (2, 3).
Men på trods af det, vi tror, vi ved, er området stadig ungt, når det gælder forståelsen af dette enorme, komplekse økosystem og dets betydning for menneskers sundhed. Den virkelighed står i skarp kontrast til "tarmsundheds"-trenden, som har været en kilde til overdrivelse og misinformation. I denne artikel ser vi på terminologien, på økosystemets opbygning, funktioner og sammensætning — og derefter på kostens rolle gennem linsen af, hvad vi ved (eller er ved at lære) om mikrobiomets indflydelse på sundhed og sygdom.
Lidt terminologi
Læser man forskning — eller bare kommentarer i medierne — om feltet, støder man uundgåeligt på en række begreber, som ikke altid bruges korrekt:
- Mikrobiom: det "udvidede genom", bakterierne leverer — altså hvilke funktioner de udøver.
- Mikrobiota: selve bakterierne i økosystemet — altså hvilke bakterier der er der, og i hvilke forhold.
- Mikrobe: mikroskopiske organismer — bakterier, arkæer, svampe, protozoer, alger og virus.
- Bakterie: encellede, meget tilpasningsdygtige organismer.
- Dysbiose: forstyrrelser i mikrobiotaens sammensætning, der påvirker sygdom.
- Præbiotikum: en selektivt fermenterbar ingrediens, der giver gavnlige ændringer i tarmmikrobiotaens sammensætning og/eller aktivitet.
- Probiotikum: en levende mikroorganisme, der i tilstrækkelig mængde giver værten en sundhedsmæssig gevinst.
- Kontrabiotikum: plantefibre, der blokerer, at potentielt sygdomsfremkaldende bakterier sætter sig fast på og trænger gennem tarmslimhinden.
Økosystemets oprindelse fortaber sig i evolutionens tåger, men flere forklaringer er blevet foreslået. Huden er udsat for luft og har gennem evolutionen været UV-eksponeret, hvilket gav bolig til iltkrævende (aerobe) bakterier (4). Tyktarmen tilbyder omvendt et mørkt, fugtigt og energirigt hjem for iltfri (anaerobe) bakterier, der selektivt fermenterer forskellige kost-substrater (4). En anden forklaring på især mikrobiotaens evne til at reagere på kostændringer er, at mennesket har koloniseret alle klodens egne og dermed levet på vidt forskellige kostformer i vidt forskellige miljøer (4, 5). Mave-tarm-kanalen er med sit overfladeareal på 250–400 m² — bortset fra huden — en af kroppens største grænseflader mod omverdenen, og den udgør både første forsvarslinje i immunforsvaret og stedet, hvor maden fordøjes og optages (5). Den enorme mangfoldighed i mikrobiotaen afspejler formentlig mangfoldigheden i miljøer og fødekilder (4, 5).
Mikrobiotaens taksonomi
Taksonomi er læren om at inddele levende organismer, og da mikrobiomet er et økosystem, kan det forstås taksonomisk. Hos mennesker er der 4 hoved-rækker (phyla) — tænk dem som militære divisioner: Firmicutes, Bacteroidetes, Actinobacteria og Proteobacteria (3). Størstedelen tilhører Firmicutes og Bacteroidetes, men vigtige populationer kommer også fra Actinobacteria og Proteobacteria (6, 7). Inden for en række findes talrige slægter, og inden for hver slægt et væld af arter.
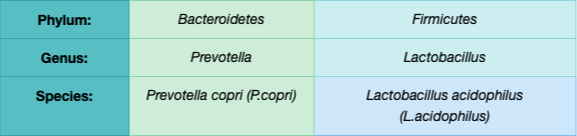
Andre økosystemer, som jord, varierer meget på række-niveau — men den menneskelige mikrobiota er bemærkelsesværdig ved sin mangel på variation her, domineret af de 4 rækker (3, 4). Til gengæld er variationen enorm på slægts- og artsniveau (3). Hvor individuelt det er, ses i en analyse af 124 nordiske og middelhavs-deltagere: der var ikke én eneste rigelig art, som to vilkårlige deltagere delte (6). Op til 160 bakteriearter blev identificeret i hver prøve, med i alt 3,3 millioner bakteriegener i hele kohorten (6).
Struktur og funktion
Hovedstedet for bakteriel kolonisering er tyktarmen (5). Der findes små mængder primært aerobe bakterier i mavesæk og tyndtarm, men de er ikke store bakterie-lokationer (5). Mavesækkens sure miljø, bugspyt- og galdesalte i tyndtarmen, ilt og den korte transittid begrænser storstilet kolonisering (8). I tyktarmen dominerer derimod anaerobe arter, specialiserede i at nedbryde bestemte kost-substrater. Visse komplekse kulhydrater passerer ufordøjet gennem tyndtarmen og bliver til fermenterbart materiale for fibernedbrydende bakterier — og sammen med den lange transittid giver det en tæt og mangfoldig bakteriekoloni (8).
Fra dette hjemsted i tyktarmen er en række roller kortlagt, som kan forklare mikrobiomets betydning for helbredet:
- modning af tarmens epitelcellelag;
- modning af immunforsvaret (tæt knyttet til ovenstående);
- regulering af inflammation;
- samspil med tarm-hjerne-aksen, der påvirker neurologiske processer og humør;
- syntese af visse vitaminer og mineraler;
- produktion af sekundære metabolitter, fx kortkædede fedtsyrer (SCFA) i tyktarmen (9–13).
Variationen i mikrobiotaen — både mellem og inden i den enkelte — afspejler et dynamisk system, der påvirkes af geografi, livsfase-faktorer (fødselsmetode og ammemetode), alder, antibiotika og kost (10).
Faktorer uden for kosten
Det har været omdiskuteret, om fostret udvikler sig i et sterilt, bakteriefrit miljø — men nyere evidens tyder på, at den første bakterie-eksponering kan ske allerede i livmoderen (14). Man har fundet et mikrobiom i moderkagen samt bakterier i mekonium (fostrets første afføring), hvilket antyder, at koloniseringen kan begynde før fødslen (14).
Ikke desto mindre er fødselsmetoden en primær bestemmende faktor for spædbarnets tidlige mikrobiota. Vaginalt fødte børn afspejler moderens vaginale sammensætning, som typisk er stabil (15). Børn født ved kejsersnit afspejler derimod hudens bakterier (hos moderen og personalet), og hudens mikrobiota er meget variabel og markant forskellig fra tarmens (15). Vaginalt fødte har rigeligt af slægterne Lactobacillus og Prevotella, mens kejsersnit-fødte har rigeligt af Staphylococcus (15). Hvor et vaginalt født barns mikrobiota begynder at ligne moderens inden for 3–7 dage, kan et kejsersnit-født barns være uroligt i op til 6 måneder (15). Der er set sammenhænge mellem kejsersnit og højere forekomst af astma, allergi og visse autoimmune tilstande — især cøliaki og type 1-diabetes (16) — men forsigtighed er nødvendig, da den slags langtidssammenhænge er svære at tilskrive én tidlig eksponering.
Ammemetoden er en anden vigtig tidlig faktor. Et særtræk ved spædbarnets mikrobiota — i modsætning til den voksnes — er, at den ikke er indrettet på mangfoldighed (17). Ammede børn er domineret af en bestemt art af slægten Bifidobacterium, der er specialiseret i at nedbryde de komplekse sukkerstoffer i modermælk (humane mælke-oligosakkarider, HMO) (17). Flaskeernærede børn har derimod højere niveauer af potentielt pro-inflammatoriske Proteobacteria og ligner mere den voksnes mønster — hvilket ikke nødvendigvis er en fordel for spædbarnets tarm, der trives på et begrænset antal specialiserede arter (17). Hos ammede bruger Bifidobacteria HMO til at befolke tarmen, udtrykke anti-inflammatoriske gener, øge immuntolerance og styrke tarmbarrieren (17). Nyere forsøg med HMO-beriget modermælkserstatning viser gavn på mikrobiel sammensætning, inflammationsmarkører og antibiotikaforbrug (18), men syntetiske oligosakkarider mangler stadig det fulde spektrum sammenlignet med modermælk (18).
Geografi former også mikrobiotaen — men den primære forskel er nok mellem traditionelle kostformer og den vestlige kost. I en analyse af befolkninger på traditionel kost i Malawi og Peru mod en amerikansk gruppe fra Philadelphia på typisk vestlig kost havde amerikanerne markant lavere mikrobiel mangfoldighed (11). Både den afrikanske og den sydamerikanske gruppe var domineret af Prevotella, der er specialiseret i at nedbryde plantefibre (11). Studiet bekræftede også, at den voksne mikrobiota udvikler sig over de første 3 leveår og derefter er relativt stabil — mens alle børn under 3 år var Bifidobacterium-dominerede (11). De tidlige faktorer er stort set bestemt for os, men de store forskelle mellem traditionel og vestlig kost peger på kostens nøglerolle (11, 19).
Hvordan kosten former mikrobiotaen
At mikrobiotaen kan skifte sammensætning som svar på kosten, afspejler menneskets evne til at reagere på ernæringsændringer på både kort og lang sigt. Forskellige bakterier er specialiserede i at fermentere forskellige substrater, så dine madvalg og hele dit kostmønster leverer råmaterialet til selektiv vækst af bestemte arter (19).
Det ses gennem hele livet. Både afrikanske børn og voksne på en traditionel kost — lav på animalsk protein og fedt, høj på fibre — har høje niveauer af Prevotella og andre arter, der kræves for at nedbryde ufordøjelige kulhydrater (20, 21). Befolkninger, der spiser rigeligt af ufordøjelige kulhydrater, har konsekvent større mikrobiel mangfoldighed end dem på vestlig kost (20, 21) — og mangfoldighed er bredt forbundet med værtens sundhed.
Disse strukturelt forskelligartede, ufordøjelige kulhydrater — resistent stivelse, ikke-stivelses-polysakkarider (NSP) og andre præbiotiske fibre — modstår fordøjelsen i tyndtarmen og når frem til tyktarmen, hvor de fermenteres selektivt (22). Et højt fiber-/NSP-indtag flytter konsekvent mikrobiotaen mod flere SCFA-producerende mikrober (20, 21). SCFA'er — især butyrat — har anti-inflammatoriske effekter i tyktarmen og er forbundet med hæmning af kræftudvikling (21).
Omvendt påvirker den vestlige kost (høj på fedt, sukker og animalsk protein, lav på fibre) mikrobiotaen negativt — og det ses allerede fra 1-års-alderen (20). I en sammenligning mellem europæiske børn på vestlig kost og afrikanske børn på traditionel kost havde de europæiske dobbelt så mange Firmicutes, færre Bacteroidetes og markant lavere SCFA-produktion (20). Den profil (flere Firmicutes, færre Bacteroidetes) er forbundet med flere pro-inflammatoriske Proteobacteria og flere sekundære galdesyre-metabolitter, der er potentielt kræftfremkaldende (23, 24).
Ændringerne ser ud til at være drevet af bestemte træk ved den vestlige kost — især fedt. Men fedttypen er vigtig. Et højt indtag af mættet fedt ændrer mikrobiotaen ved at øge galdesyre-produktionen og de galde-tolerante bakterier (21, 24). Enkeltumættet fedt ser ikke ud til at påvirke sammensætningen væsentligt (22, 26), mens omega-3 fra fiskeolie muligvis kan øge nogle gavnlige Actinobacteria og Lactobacilli og ændre galdesyre-sammensætningen — men den forskning er foreløbig (26).
Et højt indtag af animalsk protein er forbundet med øget "forrådnelse" af protein til potentielt kræftfremkaldende biprodukter, lavere SCFA og færre butyrat-producerende bakterier (24). Men protein hænger også sammen med større mangfoldighed og gavnlige forbindelser (7). Skaden ved højt protein ser ud til at afhænge af, om der er fibre med: den skete, når højt protein blev koblet med kun 8–12 g fiber om dagen (27), mens et andet studie med 31 g fiber om dagen ikke fandt øget forrådnelse eller færre SCFA trods højt protein (28). Protein-effekterne handler altså nok primært om fibre — hvilket kan forklare fordelen ved plantebaserede proteinkilder, der ofte er fiberrige (7, 22).
Mikrobiomet og tarmsygdomme
Personer med kronisk tarmbetændelse (IBD) har lav mikrobiel mangfoldighed, færre Firmicutes og Bacteroidetes og lave niveauer af butyrat (29, 30). Det ser ud til at være medieret af fiberfattige kostformer, der reducerer SCFA-producerende bakterier, svækker tarmens "tight junctions" og gør det lettere for sygdomsfremkaldende bakterier at sætte sig fast (29, 30). Netop vedhæftning af sygdomsfremkaldende E. coli er impliceret i udviklingen af IBD — især Crohns sygdom — og tyktarmskræft (31). Forsøg tyder på, at emulgatorer (allestedsnærværende i den vestlige kost) kan forstyrre samspillet mellem slimhinde og bakterier og fremme både gennemtrængning og betændelse (32).
En spirende hypotese er, at kostfibre kan forhindre E. coli i at sætte sig fast på tarmcellerne. Forsøg har vist, at NSP fra grønne pisang og broccoli blokerer E. coli-vedhæftning ved fysiologisk relevante koncentrationer (31, 33) — en forebyggende "kontrabiotisk" effekt mod en mekanisme, der er impliceret i både Crohns og tyktarmskræft (31, 33). NSP har desuden en præbiotisk effekt, der selektivt nærer Bifidobacteria og Lactobacilli og øger SCFA-produktionen samt butyrat-medieret reduktion af inflammation og kræft (25, 34).
Der er begrænset evidens for, at omega-3 kan beskytte mod IBD: fiskeolie hæmmede væksten af den sygdomsfremkaldende B. wadsworthia hos colitis-udsatte mus på fedtrig kost (29). Metaanalyser af humane data understøtter dog ikke fiskeolie mod IBD (35), selvom det at hæve omega-3:omega-6-forholdet specifikt har været effektivt til at fastholde remission (36). Samlet er data endnu utilstrækkelige til at anbefale fiskeolie til remission ved IBD (35).
Epidemiologien peger stærkt på en geografisk fordeling af IBD, der er omvendt forbundet med D-vitaminniveauer (37). D-vitamins immunologiske effekter omfatter hæmning af inflammatoriske cytokiner, øget antimikrobiel aktivitet, vedligeholdelse af tarmbarrieren og forebyggelse af dysbiose (38). Hos D-vitaminmangel-Crohns-patienter reducerede normalisering via tilskud sandsynligheden for operation markant (39), og en RCT med 1.200 IE D3 dagligt reducerede risikoen for tilbagefald ved Crohns med 50 % — hvilket, selvom det lige akkurat ikke nåede statistisk signifikans (p=0,06), var et klinisk relevant resultat (40). Ved colitis ulcerosa er evidensen for D-vitamin svag (41). Samlet peger evidensen på en gevinst ved D-vitamintilskud ved IBD, især Crohns, med et mål-niveau i blodet på 30–50 ng/mL opnået via 1.800–10.000 IE dagligt afhængigt af udgangspunktet (43).
Der er flere mekanismer, hvorigennem en vestlig kost kan påvirke risikoen for IBD og tyktarmskræft. Hos mennesker skiftede en udelukkende animalsk, mættet-fedt-rig og fiberfri kost hurtigt mikrobiotaen mod øget sekundær galdesyre-metabolisme inden for 3 dage — med hæmning af Firmicutes og Bacteroidetes og flere B. wadsworthia (24). Og det høje fedt-/lave fiberindhold øger galdesyre-produktionen, som i tyktarmen omdannes til sekundære galdesyrer forbundet med tyktarmskræft (21, 44). I et diæt-bytte-studie viste afroamerikanere på vestlig kost øget mikrobiel omdannelse af galdesyrer til kræftfremmende metabolitter, mens jævnaldrende afrikanere på fiberrig kost havde masser af SCFA-producerende bakterier og højt butyrat (21, 44). Da man byttede grupperne kost, vendte effekterne (45) — en klar illustration af sammenhængen mellem kost, mikrobiota og tyktarmskræft.
Mikrobiomet og fedme
Man har observeret forskelle i mikrobiotaen mellem slanke og svært overvægtige: sidstnævnte har flere Firmicutes og færre Bacteroidetes (45, 46). Effekterne vender ved vægttab og kalorierestriktion (47), hvilket tyder på, at ændringerne drives af kost og/eller øget fedtmasse (45).
Det er vigtigt at skelne mellem, om mikrobiomet forårsager fedme, eller om fedme afspejler kost-drevne ændringer i mikrobiomet. Ændringer som følge af vestlig kost kan øge "energihøsten" fra maden (via øget aktivitet af energi-udvindende enzymer) (7, 46), og ændret galdesyre-stofskifte kan øge fedtlagring (7, 46). Et forsøg med fækal transplantation fandt, at overvægtige modtagere øgede deres insulinfølsomhed (48), og hos personer med metabolisk syndrom gav 2 års middelhavskost mikrobiota-ændringer, der svarede til bedre blodsukker og triglycerider (47).
Men det er vigtigt at sige, at de fleste af de foreslåede mekanismer stammer fra dyremodeller, og nyere evidens tyder på, at variationen mellem individer er lige så udtalt hos slanke som hos svært overvægtige (49). Der findes til dato ingen "signatur"-sammensætning af mikrobiotaen, der kendetegner fedme (49).
Det er derfor et klart overgreb på evidensen at hævde — som nogle gør — at fedme forårsages af mikrobiomet.
Evidensen peger på, at skiftet i sammensætning er drevet af kosten — med den vestlige kost som drivkraft allerede fra barndommen (20). Fedme drives desuden af det kroniske kalorieoverskud i samme kostmønster, og de to udelukker ikke hinanden. En mere præcis beskrivelse er, at den kost-drevne ændring i bakteriesammensætning kan forværre de skadelige effekter af øget fedtmasse ved at fremme inflammation, insulinresistens og ændret fedtstofskifte (45–47).
Mikrobiomet og hjernen
Tarm-hjerne-mikrobiota-aksen omfatter centralnervesystemet, det autonome nervesystem og tarmens eget nervesystem, og kommunikationen går begge veje: hjernen påvirker tarmen, og tarmen påvirker hjernen (50). Inden for den akse er mikrobiotaen dukket op som en faktor, der kan påvirke neurologiske processer med mulige konsekvenser for humør og adfærd (50).
En mulig mekanisme er enzymet IDO, som er det hastighedsbegrænsende trin i omsætningen af aminosyren tryptofan — udgangspunktet for signalstoffet serotonin. Ved dysbiose kan høj IDO-aktivitet lede tryptofan væk fra serotonin-produktion og over i kynurenin-vejen, hvilket kan spille en rolle i både mavesymptomer ved IBS og depressive symptomer (51, 52). SCFA'er har desuden neuroaktive egenskaber ud over tarmen (53) og kan påvirke BDNF — en signalvej for nydannelse af hjerneceller, læring og hukommelse (50, 53).
Meget af det her er stadig foreløbigt, men den mest lovende evidens hos mennesker handler om stress. At indtage en fermenteret mælk med probiotiske bakterier forbedrede følelsesmæssig bearbejdning målt med hjernescanning (fMRI) (54), og en måneds probiotisk tilskud reducerede udskillelsen af kortisol i urinen — et tegn på dæmpet stressrespons (55). En mekanisme af særlig interesse er mikrobiotaens evne til at dæmpe neuroinflammation, sandsynligvis medieret af SCFA (56); at ændre tarmbakterierne har reduceret angst og depression, tilskrevet netop færre tegn på neuroinflammation (57).
Der er også antydninger af en sammenhæng med autismespektrumforstyrrelse (ASF): børn med ASF har en ændret tarmflora, og mavesymptomer korrelerer med sværhedsgraden af symptomer (58, 59). Data er dog fortsat begrænsede, og et stort randomiseret forsøg med probiotika hos børn med ASF er i gang i Italien — resultaterne afventes (60).
Hvad vi faktisk ved
På det bredeste niveau kan vi sige, at den grundlæggende kost-bestanddel, der fremmer en god mikrobiota, er kostfibre (20–26). Og blandt fibrene ser de præbiotiske ud til at være særligt gavnlige. Gibson og kolleger definerede præbiotika som en selektivt fermenterbar ingrediens, der giver gavnlige ændringer i tarmmikrobiotaens sammensætning og/eller aktivitet (61). De kendetegnes ved at modstå mavesyre, modstå enzymatisk nedbrydning og optag i tyndtarmen, og blive fermenteret selektivt med vækst af gavnlige bakterier og sundhedseffekter til følge (61).
Præbiotika findes naturligt i mad — især hvidløg, løg, hvede, havre, soja, porrer, cikorie, asparges og jordskokker (62). Til dato er kun inulin, oligofruktose (FOS) og galaktooligosakkarider (GOS) klassificeret som præbiotika med evidens for gavn (63). Det primære mål er slægten Bifidobacterium, der fermenterer de præbiotiske fibre og øger SCFA-produktionen (64). Dertil har NSP både en præbiotisk effekt og en potentiel kontrabiotisk beskyttelse mod, at sygdomsfremkaldende bakterier sætter sig fast (31, 33).
Det er værd at bemærke, at mange af de beskyttende fiber-effekter først ses ved et minimum på omkring 30 g fiber om dagen (65), og at mange af de observerede traditionelle kostformer ligger omkring 40 g — og 50–60 g, hvis man tæller resistent stivelse med (21, 44). Den optimale kost for tarmen indeholder altså et bredt udvalg af gode kulhydratkilder. Ud over fibre kan fedtets sammensætning spille en rolle: højt mættet fedt øger galdesyre og syre-tolerante bakterier (24, 29), mens en "middelhavs"-profil rig på umættet fedt kan hæmme sygdomsfremkaldende bakterier (26, 29). Endelig er samspillet mellem D-vitamin-receptoren og tarmens immunforsvar en vigtig brik: D-vitamin ser ud til at styrke immunforsvaret i tarmen og forebygge dysbiose (37, 42, 43).
Konklusion
En kost, der er høj på animalsk protein og animalsk fedt og lav på fibre, ser — på den samlede evidens — ud til at være dybt skadelig for mikrobiotaens sammensætning og funktion. Indtil der kommer evidens for det modsatte, ligger bevisbyrden hos dem, der anbefaler den slags kostformer.
Strukturelt mangfoldige, ufordøjelige kulhydrater — det, vi bredt kalder fibre — spiller en fundamental rolle i at forme en mikrobiota, der fremmer helbredet på tværs af kroppens systemer, fra tarm til hjerne. Og eftersom den voksne mikrobiota er relativt stabil, kræver det, at man fastholder kostmønsteret for at fastholde sammensætningen.
- Dit mikrobiom er reelt vigtigt for helbredet — men "tarmsundheds"-industrien løber langt foran evidensen.
- Den vigtigste kost-knap er fibre — især præbiotiske fibre og et bredt udvalg af planter (sigt højt; mange gevinster ses fra ~30 g/dag).
- Vestlig kost (meget fedt/sukker/animalsk protein, lidt fiber) forringer mangfoldigheden allerede fra barndommen.
- Skaden ved højt animalsk protein afhænger i høj grad af, om der er nok fiber med.
- Nej — fedme "forårsages" ikke af mikrobiomet. Ændringerne drives af kosten og kan forværre effekterne af øget fedtmasse.
- Tarm-hjerne-forskningen er spændende, men stadig ung: lovende på stress, men langt fra en færdig historie.
Referencer
- Sender R, Fuchs S, Milo R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016;14(8):e1002533.
- Lederberg J. Infectious History. Science. 2000;288(5464):287–93.
- Turnbaugh P, Ridaura V, Faith J, et al. The Effect of Diet on the Human Gut Microbiome: A Metagenomic Analysis in Humanized Gnotobiotic Mice. Sci Transl Med. 2009;1(6):6ra14.
- Ley R, Knight R, Gordon J. The human microbiome: eliminating the biomedical/environmental dichotomy in microbial ecology. Environ Microbiol. 2007;9(1):3–4.
- Thursby E, Juge N. Introduction to the human gut microbiota. Biochem J. 2017;474(11):1823–1836.
- Qin J, Li R, Raes J, et al. A human gut microbial gene catalog established by metagenomics sequencing. Nature. 2010;464(7285):59–65.
- Lopez-Legarrea P, Fuller N, Zulet M, et al. The influence of Mediterranean, carbohydrate and high protein diets on gut microbiota composition in the treatment of obesity and associated inflammatory state. Asia Pac J Clin Nutr. 2014;23(3):360–8.
- Donaldson G, Lee S, Mazmanian S. Gut biogeography of the bacterial microbiota. Nat Rev Microbiol. 2015;14(1):20–32.
- Kinross J, Darzi A, Nicholson J. Gut microbiome-host interactions in health and disease. Genome Med. 2011;3(3):14.
- Nicholson J, Holmes E, Kinross J, et al. Host-Gut Microbiota Metabolic Interactions. Science. 2012;336(6086):1262–1267.
- Yatsunenko T, Rey F, Manary M, et al. Human gut microbiome viewed across age and geography. Nature. 2012;486(7402):222–7.
- Natividad J, Verdu E. Modulation of intestinal barrier by intestinal microbiota: Pathological and therapeutic implications. Pharmacol Res. 2013;69(1):42–51.
- den Besten G, van Eunen K, Groen A, et al. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J Lipid Res. 2013;54(9):2325–2340.
- Gritz EC, Bhandari V. The human neonatal gut microbiome: a brief review. Front Pediatr. 2015;3:17.
- Dominguez-Bello M, Costello E, Contreras M, et al. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci USA. 2010;107(26):11971–11975.
- Neu J, Rushing J. Cesarean Versus Vaginal Delivery: Long-term Infant Outcomes and the Hygiene Hypothesis. Clin Perinatol. 2011;38(2):321–331.
- O'Sullivan A, Farver M, Smilowitz J. The Influence of Early Infant-Feeding Practices on the Intestinal Microbiome and Body Composition in Infants. Nutr Metab Insights. 2015;8(Suppl 1):1–9.
- Plaza-Díaz J, Fontana L, Gil A. Human Milk Oligosaccharides and Immune System Development. Nutrients. 2018;10(8):1038.
- Heiman M, Greenway F. A healthy gastrointestinal microbiome is dependent on dietary diversity. Mol Metab. 2016;5(5):317–320.
- De Filippo C, Cavalieri D, Di Paola M, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci USA. 2010;107(33):14691–14696.
- Ou J, Carbonero F, Zoetendal E, et al. Diet, microbiota, and microbial metabolites in colon cancer risk in rural Africans and African Americans. Am J Clin Nutr. 2013;98(1):111–120.
- Graf D, Di Cagno R, Fåk F, et al. Contribution of diet to the composition of the human gut microbiota. Microb Ecol Health Dis. 2015;26:26164.
- Walker A, Ince J, Duncan S, et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2011;5(2):220–230.
- David L, Maurice C, Carmody R, et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature. 2014;505(7484):559–563.
- Simpson H, Campbell B. Review article: dietary fibre-microbiota interactions. Aliment Pharmacol Ther. 2015;42(2):158–179.
- Singh R, Chang H, Yan D, et al. Influence of diet on the gut microbiome and implications for human health. J Transl Med. 2017;15(1):73.
- Russell W, Gratz S, Duncan S, et al. High-protein, reduced-carbohydrate weight-loss diets promote metabolite profiles likely to be detrimental to colonic health. Am J Clin Nutr. 2011;93(5):1062–1072.
- Brinkworth G, Noakes M, Clifton P, Bird A. Comparative effects of very low-carbohydrate, high-fat and high-carbohydrate, low-fat weight-loss diets on bowel habit and faecal short-chain fatty acids and bacterial populations. Br J Nutr. 2009;101(10):1493–1502.
- Devkota S, Chang E. Interactions between Diet, Bile Acid Metabolism, Gut Microbiota, and Inflammatory Bowel Diseases. Dig Dis. 2015;33(3):351–356.
- Peterson D, Frank D, Pace N, Gordon J. Metagenomic Approaches for Defining the Pathogenesis of Inflammatory Bowel Diseases. Cell Host Microbe. 2008;3(6):417–427.
- Martin H, Campbell B, Hart C, et al. Enhanced Escherichia coli adherence and invasion in Crohn's disease and colon cancer. Gastroenterology. 2004;127(1):80–93.
- Chassaing B, Koren O, Goodrich J, et al. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature. 2015;519(7541):92–96.
- Roberts C, Keita A, Duncan S, et al. Translocation of Crohn's disease Escherichia coli across M-cells: contrasting effects of soluble plant fibres and emulsifiers. Gut. 2010;59(10):1331–1339.
- Patel R, DuPont H. New Approaches for Bacteriotherapy: Prebiotics, New-Generation Probiotics, and Synbiotics. Clin Infect Dis. 2015;60(Suppl 2):S108–S121.
- Turner D, Shah P, Steinhart A, et al. Maintenance of remission in inflammatory bowel disease using omega-3 fatty acids (fish oil): A systematic review and meta-analyses. Inflamm Bowel Dis. 2011;17(1):336–345.
- Uchiyama K, Nakamura M, Odahara S, et al. N-3 polyunsaturated fatty acid diet therapy for patients with inflammatory bowel disease. Inflamm Bowel Dis. 2010;16(10):1696–1707.
- Hewison M. Vitamin D and immune function: an overview. Proc Nutr Soc. 2012;71(1):50–61.
- Clark A, Mach N. Role of Vitamin D in the Hygiene Hypothesis: The Interplay between Vitamin D, Vitamin D Receptors, Gut Microbiota, and Immune Response. Front Immunol. 2016;7:627.
- Ananthakrishnan A, Cagan A, Gainer V, et al. Normalization of Plasma 25-Hydroxy Vitamin D Is Associated with Reduced Risk of Surgery in Crohn's Disease. Inflamm Bowel Dis. 2013;19(9):1921–1927.
- Jørgensen S, Agnholt J, Glerup H, et al. Clinical trial: vitamin D3 treatment in Crohn's disease – a randomized double-blind placebo-controlled study. Aliment Pharmacol Ther. 2010;32(3):377–383.
- Nicholson I, Dalzell A, El-Matary W. Vitamin D as a therapy for colitis: A systematic review. J Crohns Colitis. 2012;6(4):405–411.
- Flanagan P, Chiewchengchol D, Wright H, et al. Killing of Escherichia coli by Crohn's Disease Monocyte-derived Macrophages and Its Enhancement by Hydroxychloroquine and Vitamin D. Inflamm Bowel Dis. 2015;21(7):1499–1510.
- Hlavaty T, Krajcovicova A, Payer J. Vitamin D Therapy in Inflammatory Bowel Diseases: Who, in What Form, and How Much? J Crohns Colitis. 2015;9(2):198–209.
- O'Keefe S, Li J, Lahti L, et al. Fat, fibre and cancer risk in African Americans and rural Africans. Nat Commun. 2015;6:6342.
- Ley R, Turnbaugh P, Klein S, Gordon J. Microbial ecology: Human gut microbes associated with obesity. Nature. 2006;444(7122):1022–1023.
- Turnbaugh P, Ley R, Mahowald M, et al. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444(7122):1027–1031.
- Ojeda P, Bobe A, Dolan K, et al. Nutritional modulation of gut microbiota — the impact on metabolic disease pathophysiology. J Nutr Biochem. 2016;28:191–200.
- Vrieze A, Van Nood E, Holleman F, et al. Transfer of Intestinal Microbiota From Lean Donors Increases Insulin Sensitivity in Individuals With Metabolic Syndrome. Gastroenterology. 2012;143(4):913–916.e7.
- Finucane MM, Sharpton TJ, Laurent TJ, Pollard KS. A taxonomic signature of obesity in the microbiome? Getting to the guts of the matter. PLoS One. 2014;9(1):e84689.
- Dinan T, Stilling R, Stanton C, Cryan J. Collective unconscious: How gut microbes shape human behavior. J Psychiatr Res. 2015;63:1–9.
- Clarke G, McKernan D, Gaszner G, et al. A Distinct Profile of Tryptophan Metabolism along the Kynurenine Pathway Downstream of Toll-Like Receptor Activation in Irritable Bowel Syndrome. Front Pharmacol. 2012;3:90.
- Clarke G, Grenham S, Scully P, et al. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol Psychiatry. 2013;18(6):666–673.
- Russell W, Hoyles L, Flint H, Dumas M. Colonic bacterial metabolites and human health. Curr Opin Microbiol. 2013;16(3):246–254.
- Tillisch K, Labus J, Kilpatrick L, et al. Consumption of Fermented Milk Product With Probiotic Modulates Brain Activity. Gastroenterology. 2013;144(7):1394–1401.e4.
- Messaoudi M, Lalonde R, Violle N, et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br J Nutr. 2011;105(5):755–764.
- Rea K, Dinan T, Cryan J. The microbiome: A key regulator of stress and neuroinflammation. Neurobiol Stress. 2016;4:23–33.
- Neufeld K, Foster J. Effects of gut microbiota on the brain: implications for psychiatry. J Psychiatry Neurosci. 2009;34(3):230–231.
- Parracho H, Bingham M, Gibson G, McCartney A. Differences between the gut microflora of children with autistic spectrum disorders and that of healthy children. J Med Microbiol. 2005;54(10):987–991.
- Williams B, Hornig M, Buie T, et al. Impaired Carbohydrate Digestion and Transport and Mucosal Dysbiosis in the Intestines of Children with Autism and Gastrointestinal Disturbances. PLoS One. 2011;6(9):e24585.
- Santocchi E, Guiducci L, Fulceri F, et al. Gut to brain interaction in Autism Spectrum Disorders: a randomized controlled trial on the role of probiotics on clinical, biochemical and neurophysiological parameters. BMC Psychiatry. 2016;16(1):183.
- Gibson G, Probert H, Van Loo J, et al. Dietary modulation of the human colonic microbiota: updating the concept of prebiotics. Nutr Res Rev. 2004;17(2):259–275.
- Van Loo J, Coussement P, De Leenheer L, et al. On the presence of Inulin and Oligofructose as natural ingredients in the western diet. Crit Rev Food Sci Nutr. 1995;35(6):525–552.
- Brownawell A, Caers W, Gibson G, et al. Prebiotics and the Health Benefits of Fiber: Current Regulatory Status, Future Research, and Goals. J Nutr. 2012;142(5):962–974.
- Bouhnik Y, Raskine L, Simoneau G, et al. The capacity of nondigestible carbohydrates to stimulate fecal bifidobacteria in healthy humans: a double-blind, randomized, placebo-controlled, parallel-group, dose-response relation study. Am J Clin Nutr. 2004;80(6):1658–1664.
- Stephen AM, Champ MM, Cloran SJ, et al. Dietary fibre in Europe: current state of knowledge on definitions, sources, recommendations, intakes and relationships to health. Nutr Res Rev. 2017;30(2):149–190.
